01/3D Structure
? About the 3D Viewer
Mol* (pronounced "molstar") is an open-source molecular visualization tool used by the Protein Data Bank and AlphaFold Database. Learn more at molstar.org.
Controls:
- Rotate: Click and drag
- Zoom: Scroll wheel or pinch
- Pan: Right-click and drag (or two-finger drag)
- Reset: Double-click to reset view
What am I looking at?
This is a predicted 3D structure of the protein. The ribbon diagram shows the protein backbone—helices appear as coils, sheets as arrows, and loops as simple lines. The shape determines how the protein functions: where it binds to other molecules, how it catalyzes reactions, and how mutations might disrupt its activity.
Color legend:
The structure is colored by pLDDT confidence score, which indicates how confident AlphaFold is in each region's predicted position:
- Blue (>90): Very high confidence
- Cyan (70-90): Confident
- Yellow (50-70): Low confidence
- Orange (<50): Very low confidence, likely disordered
02/AI Analysis
TLDR
PARKIN is a protein that acts as a quality control inspector for mitochondria (the cell's power plants), tagging damaged ones for removal—a process that fails in early-onset Parkinson's disease when PARKIN mutations occur. This AlphaFold2 structure of wild-type PARKIN shows moderate confidence (average 73%), revealing its auto-inhibited resting state where regulatory domains keep the protein inactive until needed. The structure provides a baseline for understanding how disease-causing mutations disrupt PARKIN's ability to protect dopamine-producing brain cells, which die in Parkinson's disease.
Detailed Analysis
Works Cited
Similar Research
03/Research Data
ClinVar Classification
Not found in ClinVar
Population Frequency
No population data available
Disease Associations
947 totalShowing 5 of 947 associations
AI Research Brief
Research brief will be generated when agent findings are available.
04/AlphaFold Metrics
05/Domain Annotations
Structural Domains & Regions
Functional Sites
Binding Partners
Gene Ontology
06/Structural Caption
AlphaFold2 structure of wild-type PARKIN (O60260) shows well-folded ubiquitin-like and RING-IBR-RING domains with flexible linker regions, consistent with auto-inhibited state.
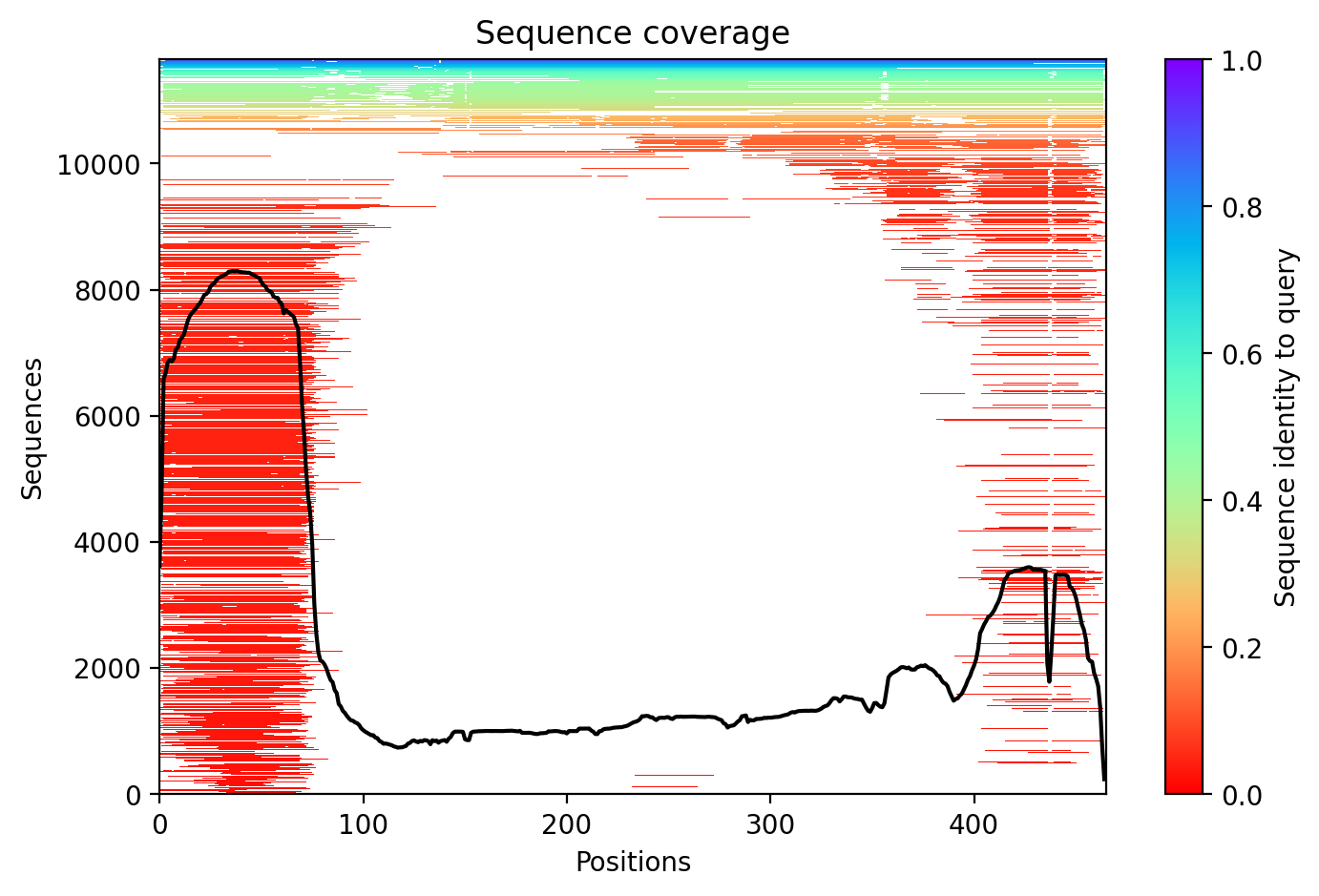
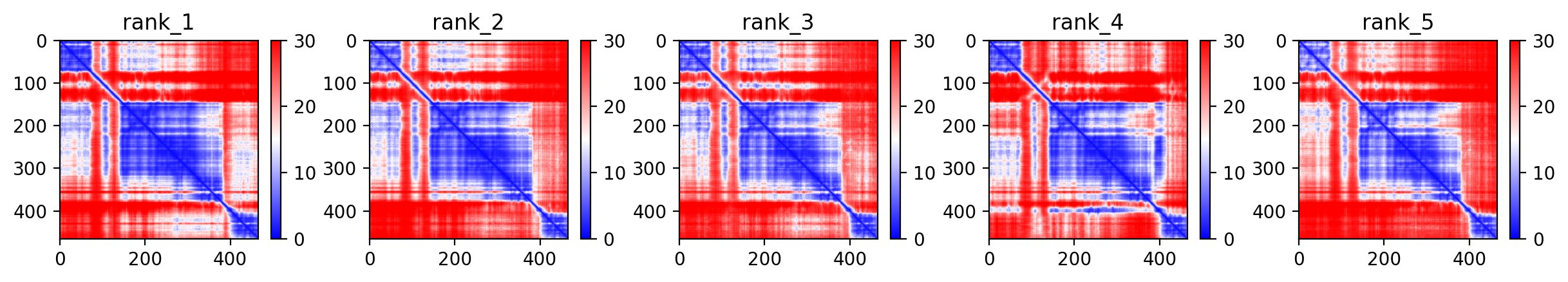
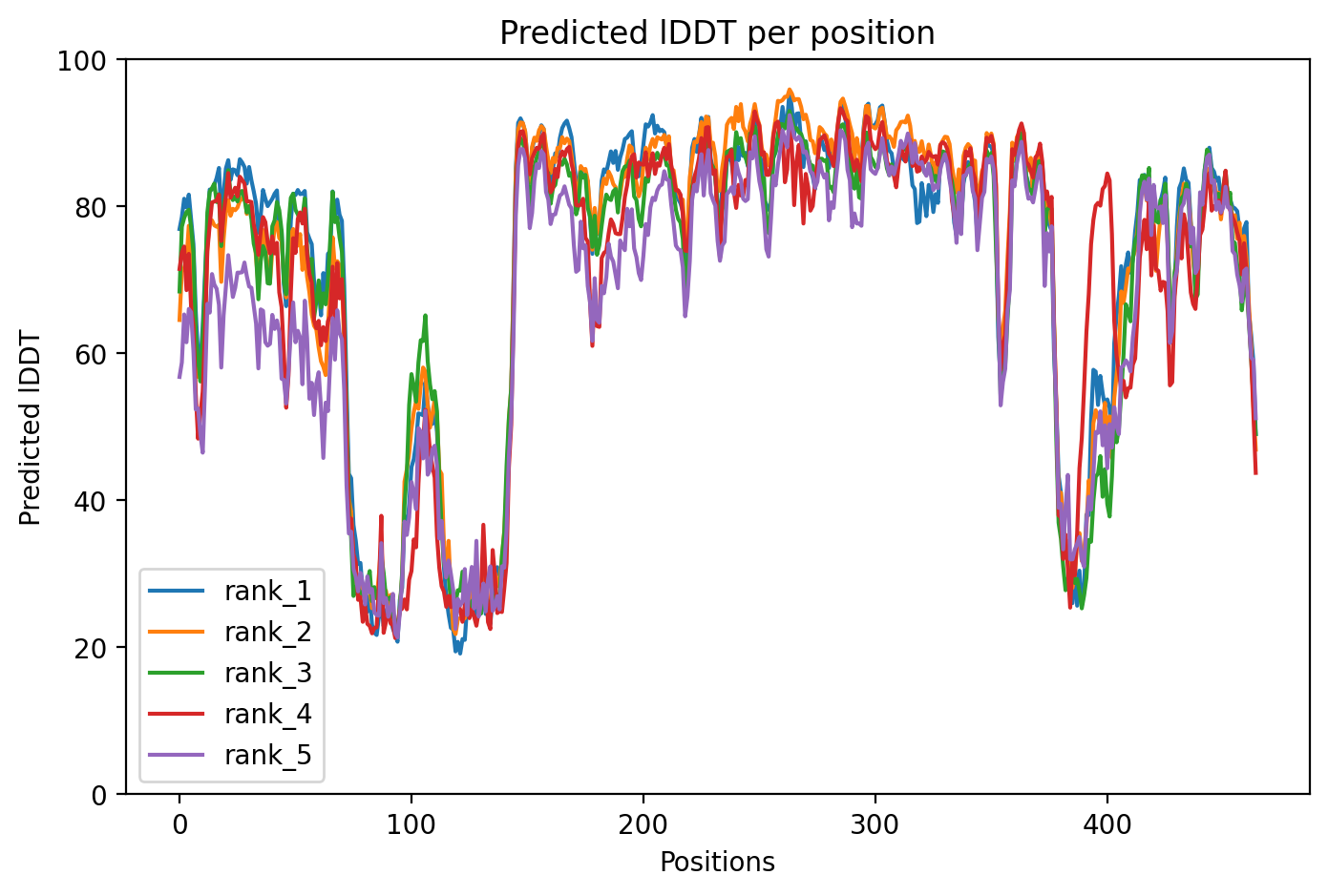
Average pLDDT of 72.7 with 71% high-confidence residues indicates moderately well-folded structure. The N-terminal ubiquitin-like domain (residues 1-76) and structured RING/IBR domains show higher confidence, while linker regions including the disordered segment (residues 77-99) exhibit lower confidence scores.
High-confidence regions map to the functional ubiquitin-like domain and the catalytic TRIAD supradomain (residues 234-465) containing RING1, IBR, and RING2 domains. The disordered linker region (residues 77-99) and interdomain connectors show expected lower confidence, consistent with their flexible nature in the auto-inhibited PARKIN structure.
Wild-type fold — no variant mutation. This represents the native auto-inhibited conformation of PARKIN E3 ubiquitin ligase.
07/Peptide Therapeutics
Aggregation analysis pending. Run peptide agent to compute aggregation propensity.
08/Known Inhibitors
No known inhibitors found. Run peptide agent to search literature.
09/Candidate Peptides
No candidate peptides generated yet. Run peptide agent to design inhibitory peptides.
10/Agent Findings
No agent findings yet. Research agents analyze folds on scheduled intervals.
11/Agent Annotations
No agent annotations yet. Agents can submit annotations via the API.